Francisco Torrent-Guasp
HEART STRUCTURE AND FUNCTION - FUTURE DIRECTIONS
![]()
![]()
![]()
![]()
|
|
Francisco Torrent-Guasp HEART STRUCTURE AND FUNCTION - FUTURE DIRECTIONS |
|
|
|
|
To see the figures bellow enlarged, please just click on the figure´s miniature.
To come back again, please press "BACK" button on your browser. |
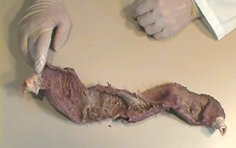
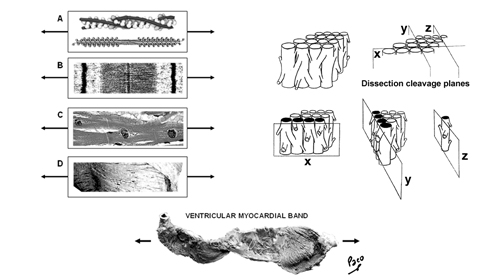
You may download this page as Adobe Acrobat PDF file (237 KB) following the link bellow. Toward New Understanding of The Heart Structure and Function "Hos igitur quasi vestigiis odorati tradendae medicinae initium ab humano corpore ducemus, quod et artis subjectum existit, et omnium primum sensibus occurrit notissimum: a quo dein per minima quaeque deducti ad ea denique mentis impulsu feremur, quae cogitatione sola comprehendi possunt" "We shall start the beginning of the teaching of medicine from the human body, which is both the subject of the art of medicine and, first of all, it comes most clearly under our senses. Then from there, led through all the minutiae, we shall be finally carried by an impulse of the mind, to those things which can be understood by thinking alone." Jean Fernel (1497-1558), physician to King Henri II of France Toward New Understanding of The Heart Structure and Function Introduction One of the most important scientific missions in this century is integration of basic research with clinical medicine. With substantial achievements at genetic, molecular and cellular levels, during the past few decades, recent advances in elucidating myocardial stucture and function have made a paradigm shift in research and provided a promising ground for the new integrative knowledge of the heart structure and function. The ancient enigma of myocardial architecture is finally solved. Discovery of the ventricular myocardial band (VMB, Figure 1) revealed unavoidable coherence and mutual coupling of form and function in the ventricular myocardium, urging for reconciliation of some exceeded concepts about electrical, mechanical and energetical events in human heart. Careful integration of this knowledge is not of merely academic importance, but is also the essential prerequisite in clinical evaluation and treatment of different heart diseases. The best example are recent modifications and enrichments in heart failure conceptions, leading to new therapies addressed to “disease and not to symptoms”. Kresh and Armour advised that “heart should be considered as self-regulating functional system, which is greater than the sum of its constitutive parts”. This general statement should be adopted as a common standpoint for those, who intend to participate and contribute in integrative heart researches. Today, more and more, parts of the road to excellence are leading to collaboration between the scientists from quite different branches. Many of them, from the mathematicians to the cardiac surgeons, are already discussing about heart, in very comprehensive manner. But even so, there are still many points of missunderstandings and disagreements. Most of them are emanating from different research technologies applied, while some others, are still difficult to explain. To overcome these diversities, we should first be aware that scientific theories are validated by empirical testing against physical observations, rather than by simple judging of their logical compatibility with the available data. But, when we reach the point upon which our scientifical perception is substantiated, as Fernel suggests, "we shall be finally carried by an impulse of the mind, to those things which can be understood by thinking alone". Thus, in order to fully understand complex three-dimensional architecture of the ventricular myocardium, some essential and scientifically validated facts should be summarized and emphasized again. Spatial organization of the ventricular myocardial fibres – historical scope The problem of the macroscopic structure of the ventricular myocardium has remained unsolved since the XVI century, when anatomy became an authentic science. The spatial organization of the myocardial fibres has been represented, as James Bell Pettigrew wrote in 1864: "an arrangement so unusual and perplexing, that it has long been considered as forming a kind of Gordian knot of Anatomy. Of the complexity of the arrangement I need not speak further than to say that Vesalius, Haller, and DeBlainville, all confessed their inability to unravel it." Reviewing the scientific reports, made along the last five centuries, for those who have made a substantial contribution to the knowledge of rule-based assignment of different trajectories followed in the space by each ventricular myocardial fibre, one realizes that only Richard Lower’s work, reported in 1669, provides still irremovable anatomical fact. He describes that, in the left ventricular wall, two groups of fibers could be distinguished. Superfitial fibers, that are coming from the base and run subepicardically, suffering a reflection at the apex, after which, they become subendocardial, coming back toward the base. The second group (i.e deep fibers), are located in the middle of the ventricular wall thickness, and run in a progressive transverse fashion. All successive contributions, including those made in the first half of XX century, more or less, limit themselves to repeat, although in different terms, these Lower's findings. Thus, in 1749, Senac arrives to the conclusion that the outer fibres, corresponding to the external and internal surfaces of the ventricular wall, run in an almost vertical direction, meanwhile, the inner ones are extended circularly in a near horizontal plane. Later on, in 1792, Wolf observes that within the deep fibres could be distinguished two trajectories. The external one - along which deep fibers run progresively more near to the direction followed by the subepicardial fibres, and the internal one - progressively inclining toward the subendocardial ones. In 1823, Gerdy, also according to the description of the English pioneer[15], claims that the inner fibres of the ventricular wall, coming from the ventricular base, fit with uninterrupted succession into the outer, subepicardial and subendocardial fibres, making an opened figure of eight. Weber states once more, in 1831, that the more superficial fibres, (i.e. subepicardical and subendocardical), coming from the ventricular apex, irradiate to arrive the ventricular base. Ludwig has noted, in 1849, that uninterrupted fibers are forming a closed figure of eight while passing around left ventricle, changing their angle smoothly from the epicardium to the endocardium. In his comprehensive study, published in 1863, Pettigrew establishes the existence of seven distinct muscular layers, that can be individualized by the progressive change in direction of their fribres, from the epicardium to the endocardium. Krehl conceptualized in 1891 the “Triebwerk”, as nested set of fiber paths, describing a subepicardial and subendocardial continuity at the level of the mitral orifice, and at the apex of the left ventricle, where they invaginate. MacCallum in 1900 and later on his teacher Mall[23] in 1911, described separately, again according to Lower, that the ventricular wall is made of fibres, with an “V” configuration, which adapt progressively one into the other, getting more and more opened “V” shape, while going to the middle regions of the ventricular wall. Mall also describes two big muscular fascicles, “sinospiral” and “bulbospiral”, in their turn divided in superficial and deep, but honestly underlying that he can not give any simple schema “which applies equally well to all portions of the ventricular wall”. Later on, in 1956, Lev and Simkins described three different fascicles: one epicardial, other endocardial and a third one, located at the middel of the ventricular wall. Lev, according to Mall, claims that “the exact explanation for the direction of the various fasciculi, depends upon comprehensive embriologic studies of the myocardium, which are still to be carried out”. Hort has made, in 1957, an important contribution with “micrometric evidence of shifting interdigitations of individual neighbouring fibres”. Daniel Denison Streeter, with his important works from 1966 to 1979, undoubtedly deserves the attribute of "crossroad" between the classical and the modern approaches in studying the heart structure and function relationships. With unique ability to “create a picture from the puzzles”, he joined his “opened-up Japanese fan” concept of the progressive change of direction in the successive transmural planes of fibres, with reproducible methodology described by Hort and Torrent-Guasp into complete mathematical study and comprehensive description of geodesic trajectories of the ventricular myocardial fibres. The result of anatomical studies, giving rise to the VMB concept, provided that simple schema, about which was claiming Mall, that applies equally well to all the ventricular myocardial fibres, showing them joined together in a coherent common general architectural plan. As emphasized by Streeter, “accepting the statistical criterion of the predominant fiber direction at given point”, earlier described by Torrent-Guasp, accomodates factual difficulties, arising from complex and anisotropic myocardial architectural design. Therefore, in the ventricular myocardium, there are no anatomically reproducible muscular fibres or distinctive layers, as thaught by predecessors or erroneously interpreted by some contemporary researchers, but only consistent vectorial functional trajectories and cleavage plans, which could be very easily reproduced by previously described dissection method (Video 1).
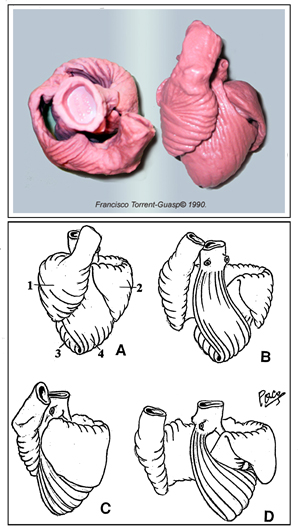
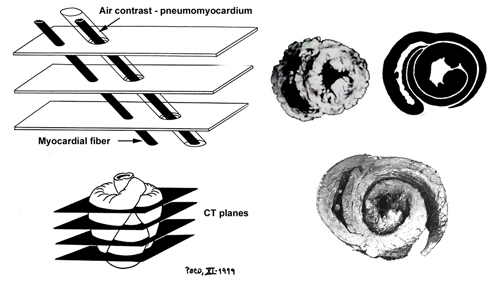
Video 1 (rm file 862KB - Real Audio Player) The knowledge of that morphology, which configurates a helicoid with two spiral turns, has been the base that has led to achieve the explanation of the way the heart performs its double function, ejection and suction of blood, a problem arised 2300 years ago by the Greek physician Erasistratus of Chios (304-250 BC). Harmony of form and function From the earliest days of anatomy and physiology, the form and the function were considered to be an inseparable attributes of a whole, either form teleological or from mechanistic (i.e materialistic) points of view. It is simply not possible to understand the essence and the magnitude of natural harmony between form and function, unless we accept the advice given by Sommer and Johnson that "relating structure to function in any organ, ...leads inevitably to ontogeny and phylogeny" and unless we expand our intelectual scope from the molecular to the organ level. Recent brilliant review on cardiogenesis by Moorman and Christoffels, integrates complex mechanisms, involved in transition from a peristaltic tubular heart to a synchronously contracting four-chambered organ. Development and patterning of vertebrate heart is amenable to different, evolutionary conserved and specific transcriptional networks, as well as to the variety of epigenetical influences. New “balooning model of chamber formation” offers a logical explanation, from the genetical and functional points of view, of the four-chambered heart design and the organization of its electrical acitivity, thus overcoming the mayor week points of previous ”linear array” concept, being considered as “one of the most fatal assumptions”, regarding morphology and flow direction. Having on mind that evolution does not “conserve” phenotype but genotype[39], it is easy to understand that phylogenetic memory does not neccessairly produce morphological equivalents during ontogenesis, but rather appears in a form of “fast-running” genetic programes. Accordingly, Ernst Haeckel's biogenetic law, stating that: “ontogeny is the short and rapid recapitulation of phylogeny”, does not necessarily mean that ancestral morphology must appear in a recognizable manner during embriogenesis. Phylogenetic and ontogenetic analyses and comparisons, should take into account a fact, that patterning, rather than structural appearance, is the only relevant and potentially accessible information. Specifical spatial changes of the heart tube and surrounding blood vessels are consequences of genetically programed proliferative and apoptotic events, as well as epigentically induced remodelings. Primordial cells originating from primary and secondary heart fields, as well as those from neural crest and proeipcardium - all participate in a final composition of the heart. Epithelial-to-mesenchimal transformation of the endothelial layer in a developing heart, is a nice example of biologically and evolutionary “condensed genetical knowledge”. Endothelial cells, underwent such transformation, start a series of mitotical divisions, filling the acellular “cardiac jelly” and producing the visible protuberances denominated as endocardial cushions and trabeculae. Similar process occurs on the other side of cardiac jelly, in a compact myocardial layer, without previous epithelial-to-mesenchimal transformation. Interconnections of those cellular bridges, surrounded by cardiac jelly, define the final spiral patterning of the adult ventricular myocardium (VMB). Transepicardial (proepicardial) proliferation supports the myocardial mass by vasculature, along with cardiac and noncardiac (mesocardial, neural crest) connective tissue, giving the rise to extracellular matrix, insulating tissues and anchoring fibrous structures within the heart. Filling the cardiac jelly with cells (i.e. “compaction of the ventricular wall”) is, maybe, the most important mechanism, that governs the specific transmural spiral patterning of the ventricular myocardial fibers, resulting in formation of the double helical VMB. Accompanying visible structural changes (genetiaclly and epigenetically controlled), so often described as a distinctive phases of embriological cardiac modeling, are probably notihing else, but our “snap-shots”of this continuing patterning process. Looking back, into predecessors' circulatory systems, genetically, morphologically and functionally, we could not resist to conclude that pumping organs (hearts) have been developed later than tubular, vascular system. Moreover, it is evident and emphasized in our previous reports, that specialized pumping chambers have been developed from the tubular vascular system. Shigei et al, have nicely reviewed phylogenetical and ontogenetical changes within tubular vascular system of invertebrates and vertebrates. According to them, the appearance and development of the “endothelium or endothelial tubular system” and “sympathetic nerve/medial smooth muscle system”, were respectively, two most remarkable changes that have occurred in the course of evolution of vertebrate circulation. They hypothesize that the sympathetic nerve/medial smooth muscle system, may be regarded as a new neuroeffector mechanism, developed for systemic regulation of the endothelium-lined closed vascular system. The complexity and difficulties in maintaining the homeostasis, have urged for development of more sophisticated control mechanisms, which may sometimes overwhelm intrinsic functional capacities in circulatory system. Accordingly, both the heart and the tubular part of circulatory system (i.e. arteries, capillaries, veins), now being separated in pulmonary and systemic circulation, have been submitted to numerous adjustments (e.g. complex neural, humoral, rheological and other influences) during phylogenesis and ontogenesis. Structural features of each constitutive component of the heart, reflect both its electrophysiological and elastomechanical performances, et vice versa. Manasek has shown that longitudinal re-arrangement of developing myofibrils and appropriate re-shaping of primitive cardiac myocytes, are induced by changes in haemodynamic forces. Micro-design of a single cardiac myocyte is, thus, amenable to fine structural adjustments, in order to retain the optimal functional capacity in a different loading conditions. Gregorioy has proved that step-like sarcomeral organization is necessary for synchronized electro-mechanical action, which produces the unidirectional blood flow in a hearts of all amniotic animals. Accordingly, size, shape, connections and predominant orientation (in a three-dimensional space) of a single-cell sarcomeral protein molecules, determine their functional behaviour. The same is true for each individual myocardial cell, myocardial fiber (being a series of longitudinally and laterally connected cells), and myocardial laminar sheet (Figure 2). Microscopical and macroscopical myocardial architeture, and particularly the existence of myocardial laminae or fiber bundles, separable by distinct anatomic cleavage planes, have been a controversial subject, since long ago. LeGrice et al, however, have provided acceptable solution, documented by their comprehensive, detailed measurements of canine ventricular myocardium. They have reported that the myofibers are arranged into distinct myocardial laminae, three to four myocytes thick, separated from adjacent laminae by the extracellular collagen network. The myocytes are tightly coupled within the laminae but sparsely coupled between adjacent laminae. The planes of the laminae could be defined locally by the longitudinal axis of comprising myofibers and by their spiral transmural direction on the ventricular mass level. Integrative knowledge of rule-based assignments, on different organizational levels within ventricular myocardium (Figure 2), is essential prerequisite for understanding that blunt anatomical dissections, following predominant fiber direction, are revealing their unique functional rather than eclectic anatomical personality. In addition, understanding of this principle would, hopefully, prevent erroneous comparisons of the VMB with skeletal muscle model, and enable those with individual difficulties to follow described VMB blunt dissection protocol (Video 1), to adopt this easy-to-learn technique (Figure 3). Since the mechanical result of the contraction of any muscle, always depends of its disposition in the space - the direction defined by such muscle, will have the gratest influence on its particular action, when being activated. This interdependence of myocardial form and function, described in VMB concept, has been recently validated on intact, beating hearts, in numerous investigations. The first, indirect visualisation of the VMB fiber trajectories in intact bovine heart, was done by Lunkenheimer et al. Using a series of computerized tomographies, after transcoronary produced pneumo-myocardium (Figure 4), he was able to trace directly only the connective tissue scaffold of the ventricular musculature, arranged in a series of differently orientated spirals. Although this, basically, double-contrast technique, was not able to visualize myocardial muscular compartment (due to its dessication and partial fragmentation by the pressurized interstitial air), it has left us a solid “negative image” of its spatial orientation, which, as the author said: ”…confirms and extends Torrent-Guasp’s double loop concept”. Recent advances in imaging techiques, have provided even more reliable methods for myocadial structural and functional analyses. Material anisotropy of living tissue has been mapped using diffusion-tensor magnetic resonance imaging (DTMRI). It has been established that water diffusion anisotropy, measured by MRI, faithfully parallels histologic anisotropy. DTMRI studies on intact, human beating heart, have confirmed that myocardial fibers within the ventricular mass, are arranged in layers of counter-wound helices, encircling the ventricular cavities. Displacement encoding stimulated echo (DENSE) MRI, provides a high spatial density of displacement measurements in the myocardium, via stimulated echoes, while the image is always acquired at the same time point in the cardiac cycle (Figure 5). The spatial and temporal resolution of the data acquired using this technique is superior to any other imaging modality involving myocardial tagging and tracking. In recent report, Saber et al., explains that it was possible to determine both myocardial fiber orientations and ventricular wall dynamics in vivo, based on the VMB concept, by applying DENSE-MRI technique. Many lines of evidence suggest that this fiber configuration serves to equalize stresses and strains across the thick-walled ventricle, allowing both active and passive tissue components to operate in optimal mechanical regimes during the cardiac cycle. Horovitz et al., claimed that: “Anatomical findings as well as theoretical considerations indicate that the myocardial fibers lie along minimal length geodesics of the left ventricular wall.” It is also a well known fact, that myocardial cells are optimized to shorten along their long axis. In a true physiologic situation, the radius of curvature, sarcomere length, fiber orientation, wall thickness, and electrical activation vary widely with location within the ventricular wall and within different time frames, during the cardiac cycle. Even so, the ventricles, being considered as remarkable product of natural engineering, have the unique ability to translate 15% linear sarcomere shortening into ejection fractions of greater than 50% and wall thickening greater than 30%. Such high efficiency coefficient could not be explained without taking into account a specific three-dimensional architectural plan, explained in VMB concept. Unraveling helicoid, configurated by VMB, allow us to distinguish four segments whose respective fibers adopt different directions within left ventricular mass. These four segments, when succesively actvated during the cardiac cycle, can be further functionally individualized, through the different actions they perform during the contraction. Those actions, as explained in our previous reports, provide the healthy heart with strong capacities (defined by specific spatial arrangement of the VMB segments) to ensure its own efficient emptying (systolic function) and filling (diastolic function), by means of successive muscular contraction along the VMB. Furthermore, the true understanding of the VMB biomechanics, allows clear insight in the venricular torsional mechanics and systolic wall thickening, a phenomena clearly visible by different imaging techniques, but hardly explainable by classical knowledge of ventricular structure and function. Myocardial fiber architecture is also a key determinant of both normal and pathological electrical properties of the myocardium, strongly influencing initiation and spread of the ventricular dysrhythmia. Along with the mechanical aspects of the ventricular performance, attention has been refocused again, on the electrical events, giving rise to excitation-contraction coupling along the VMB. Those events, and particularly their anatomical and molecular substrates, still remain controversial. Examining elctrophysiological and functional data, in order to test the hypothesis of activation sequence within the ventricular myocardium, James Cox has calculated the delivery of the impulse throughout the VMB. He had carefully plotted out how the electrical impulse was delivered to the myocardium, by the specialized conduction system, and correlated those findings with the velocities of conduction in thin and thick areas of the heart. It came out that impulse delivery precisely mimicked predicted sequence of activation. It has to be emphasized here, that patterns of contractile activities within ventricular myocardium (i.e. impulse deliveries), may or may not precisely correspond with patterns of electrical excitations, as recorded by microlectrodes or voltage sensitive dyes. This fact is essential for the explanation of different results, obtained by ventricular electrical mapping studies, and those that are analyzing sequences contractile activities within ventricular mass. It has been clearly demonstrated by fast Fourier analyses of ventricular MUGA scans, that muscular contractile activity, during the cardiac cycle, progresses (in a peristaltoid manner), along succesive VMB segments. Since the excitation necessairly preceedes contraction, the most logical pattern of ventricular electrical activation should follow, both spatially and temporarily, previously descirbed sequence of its mechanical action. Recent analyses of temporal shortening at the sites of sonomicrometer crystals, implanted in selected regions of intact animal heart, have validated proposed sequence of contraction along the VMB. Apart from initial encouraging results, more studies are needed to confirm that impulse delivery (i.e. excitation-contraction coupling), is following the sequence of activation along the VMB. Current anatomical and electrophysiological knowledge neither fully accept nor deny this pattern of electro-mechanical coupling within ventricular myocardium. The most controversial topics are related to the organization and function of, so-called, “specialized conduction system” within heart. There have been numerous attempts to explain the origin, development, organization and function of the conduction system, both in health and in disease. It is evident fact, from the pertinent literature, that cardiac myocytes, conventionally distinguished as working myocardium and specialized conducting system, share the same embriological origin. Phylogenetic and ontogenetic studies have shown the early appearance of the unidirectional, slow peristaltic waves of contractions along the heart tube, prior to any distinguishable molecular or other phenotypical differentiation of existing myocites. Patten concluded long ago, from his experimental studies that: “the whole of the primary myocardium, constituting the wall of the myocardial tube, was acting as a conducting tissue.” Recent reports on fetal myocite transplantation, and consecutive reestablishment of myocardial pacemaking activity, seems to be in accordance with previous statement. During the further developmental stages, it appears that certain myocite populations maturate faster than the others, giving a rise to the anisotropy of conduction velocities, which is, on the other hand, necessary for coordinated activation of different segments within developing heart. This maturation, among other aspects, was shown to be related to specific ion chanells, and paradoxically, those cells that we call “specialized” are in fact less mature than cells belonging to the working myocardium. Pacemaker activity, while present in embrionic ventricular myocytes is lost in adult ventricular myocytes because it is normally held back or repressed by the presence of inward-rectifier potassium chanells. Inhibiting those chanells in working cardiac myocyte, by adenoviral gene transfer, Miake et al., were able to produce a cells with higher spontaneous depolarization rates. There were many attempts to define some universial conductive genotype and phenotype, but this task still remains to be accomplished. We find it interesting to take into consideration some epigenetical influences, which may play a certain role. Since the electrical propagation through the developing heart (being basically physical event) follows the pathway of the lowest resistance (according to Ohm’s law), than the cumulative electromotive force, could be dependent on spatial organization of myocardial cells. In other words, the structural patternning of the developing heart, may have an influence on further functional (and structural) differentiations toward slow and fast conducting compartments. From the laboratory to bedside To close the circle, the new morpho-functional concept of the VMB has to be widely integrated in clinical practice. In its present extent, the VMB concept does offer solutions to some ongoing clinical controversies. During the past two decades, a new branch of cardiology has been developed, on the basis of scientifical efforts to understand, define and treat diastolic heart failure. As explained in our previous work, the concept of active diastole has been offered a time ago as possible solution. Untill detailed explanation of VMB biomechanics was available, it was hard to undestand the phenomenon of the active ventricular filling. Contemporary experimental and clinical investigations, unequivocally support the attitude that only powerful suction force, developed by the normal ventricles, could be able to produce an efficient filling of the ventricular cavities. The most important part of ventricular diastole (i.e. the rapid filling phase), during which it receives more than 70% of the stroke volume, belongs to the active muscular contraction of the VMB ascendent segment. Therefore, its of utmost importance for the clinical practice to realize, that normal ventricular myocardium possesses strong intrinsinc capacities (due to specific spatial arrangement of the VMB segments), to ensure its own efficient emptying (systolic function) and filling (diastolic function), by means of successive muscular contraction along the VMB. By understanding this principle, there would be no longer doubts, whether there is: “Diastolic heart failure or heart failure caused by subtle left ventricular systolic dysfunction?. As Yip et al., concluded:” time for a redefinition” has come. We are all aware that hallmark of this century is an increasing number of patients with hart failure. A lot of them will need some kind of surgical treatment. So far, several surgical ventricular restorative procedures were developed on the basis of VMB concept. Vaage has nicely concluded in his recent editorial, that basic science “helps and promote the development of cardothoracic surgery as a modern speciality with a dynamic and scientific profile”. Therefore, instead of empirical, we should try to develop new surgical procedures using more rational approach. Trying to find an optimal non-transplant options, many surgeons were trapped in purely mathematical evaluation of their surgical strategies. These “Laplacian” procedures (although some of them being very elegant), are commonly neglecting some essentially biological principles (e.g. tissue composition, viability, inflammation), governing behaviour of both normal and diseasaed heart. To develop more physiological heart failure surgery - the first thing that we have to realize is that the heart is not a “soap bubble”. In another word - to deal with a heart failure - we have to understand the function of the normal heart. To do so - we have to understand its structure. Apical loop of the VMB (Figure 6) is the principal motor of the heart. Cutting through this loop, as Batista proposes, not only reduces the left ventricular volume, but impairs both its systolic and particularly diastolic performance. But if we preserve the apex, as we have suggested long ago, and as Komeda et al., have done in large experimental animals, the mayor disadvantages of the left ventricular reduction surgery could be avoided. Understanding of the VMB concept is of critical importance in performing a variety of restorative surgical procedures, all of them addressed toward unfavourable left ventricular remodeling. We reasonably expect to see a lot of new clinical implementations in the nearest future, as already seen for congenital heart defects. Important research directions The VMB concept has emerged from more than 50 years long research with more than 1000 anatomical dissections of the hearts, belonging to different species. Although the first publication with detailed anatomical study has appeared more than 20 years ago, a serious attention of the wider scientifical community was not evident, untill NIH and NHLBI have organized the workshop: “Form and Function: New Views on Disease and Therapy for the Heart” (Bethesda, MD, 2002). Since then, this new anatomical and functional concept has become seriously considered in many researches, but still, there are many questions to be answered. The architectural plan of the ventricular myocardial fibers, represented by VMB, which describes two spirals in the space during its trajectory from the pulmonary artery to the aorta, defining a helicoid which delimitates two ventricular cavities, rises many interesting questions related to phylogeny and ontogeny of the heart. In spite of perplexing amount of informations, obtained from contemporary molecular and genetical analyses, it remains unclear the true origin of evident anatomical, histological and functional differences between atria and ventricles of the heart (Figure 7), on the one side, and the apparent histological and functional similarities between veins and atria, or between arteries and ventricles, on the other side. The form and the function of the heart (as well as of any other structure) are inevitably interdependent. The same, we believe, is true at each phylogenetical and ontogenetical level. Regarding previous, one of the most challenging question is realted to the transition from the aquatic to the terrestrial breathing. Precisely, the only pumping chamber in the circulatory system of the pisces belongs to the venous part (bringing the venous blood to the gills). The absence of pumping chamber in the arterial part of their circulation, is explained by low metabolic rates (low resistance to blood flow) and the "whole body movements", both being suficient for adequate blood supply of the systemic capillary network. This example rises several concerns. First, is it really low metabolic demand and whole body movement a consistent explanation for the "un-pumped" blood flow in the arterial system of the pisces or, maybe, a kind of vestigial vascular peristatic waves may be the additional blood propeling force? Second, concerning the function (pumping the venous blood toward the gills), the entire heart of the pisces is in fact the venous pump. Morphologically, it looks like a heart tube of the higher, terrestrial breathing species (aves, mammals). This similarity in morphology, with obviously different (more complex) function is confusing. Available explanations (more or less) could offer some insight into morphological transformation of the serially connected chambers within heart tube - into parallely connected chambers of the final organ. From the functional point of view – it remains unclear how does the segmented organ which pumps exclusively venous blood (i.e. thw heart of the pisces) - acquires anatomical and functional connection with the systemic, arterial circulation in higher species, having in mind that its entire “arterial pole” (i.e. aortic sac) is actually deriving venous blood to the gills? Phylogenetical and ontogenetical studies and meta-analyses, performed along with VMB researches, have opened completely new fields, now being conceptualized in several ongoing research pojects. One of them is related to electrical activity of the heart and blood vessels and is supposed to analyze possible presence of the vestigial vascular peristatic acitivity. It is widely accepted that electrical coupling between myocardial cells is acomplished by cell-to-cell transmission, via the gap juctions. Recent advances in understanding the subtile mechanisms of spreading electrical activation, by proton (H+) hopping between hydrogen bonded water molecules have stressed the importance of the interstitial tissue composition in that process. Changes in the electrical field, transmitted by Purkinje network, may alter the ionization of water molecules bonded to the matrix proteoglycans and this may, in turn, produce successive proton hopping along those molecules. Such kind of monovalent cation mobility is remarkably faster than Na+ or K+ diffusion, responsible for cell-to-cell electrical conduction. Having in mind again, that electrical activation preceedes the mechanical contraction, it has been observed that the earliest contractions in human heart appear 35msec after initial excitation, and that most of the fibers are entering contraction within 105msec. Taking into account the 98% limit of the QRS duration of 116msec in normal human subject, and the maximal possible velocity of the cell-to-cell conduction of 0.3mm/msec (along the longitudinal axes), it comes out that maximal lenght of the unraveled VMB should not be greater than 3.5cm. In a reality, just as a comparison, the average lenght of its ascending segment (in adult human heart) is 13cm. Therefore, proton hopping theory of electrical conduction, may bring some new light on excitation-contraction coupling at any given heart rate. The importance of the mitral apparatus to left ventricular function has been suggested in many clinical and experimental studies of mitral valve replacement. True reasons for better ventricular function with preserved “annulo-papillary continuity” still remain unclear. Development of new experimental research technolgies has allowed us to design an experimental protocol which would (hopefully) prove our thesis about the role of the papillary muscles and chordae tendineae within the mitral aparatus and their influence on the ventricular function. Finally, based on VMB anatomical and functional concept we have designed an external cardiac supporting device (“brace prosthesis”), which is ment to preserve both systolic and diastolic functions of the heart (Figure 8) and prevent a series of events accompanying process of ventricular remodeling. This project is currently in the experimental phase, and we hope that we would be able to report some results in a nearest future. Conclusions As Streeter said: “No man-made structure is designed like a heart. Considering the highly sophisticated engineering evidenced in the heart, it is not surprising that our understanding of it comes so slowly.”. The VMB concept, developed over the last 50 years, has gathered many people willing to contribute to the knowledge of heart structure and function. All of them, led by the misterious call of unkown, are still aware that our mutual final goal is yet to be achieved. Whether “the helix and the heart affect nature, the heart, and the human”, we really don’t know, but one thing is evident – we are still together on the road toward new understanding of the heart structure and function. |
|||
|
Figure 1 |
||||
|
Figure 2 |
||||
|
Figure 3 |
||||
|
Figure 4 |
||||
|
Figure 5 |
||||
|
Figure 6 |
||||
|
Figure 7 |
||||
|
Figure 8 |
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
References:
|
||||
|
||||